TRABAJOS
Sistema Silvopastoril con diferentes coberturas arbóreas habilitado por
rolado de baja intensidad. Impacto en glomalinas y fracciones de carbono
del suelo
Analía Liliana Anriquez1; Susana Arias1; Juan Eduardo Silberman1-2; José Alfonso Domínguez
Núñez3; Carlos Guillermo Kunst4 & Ada Susana Albanesi1*
1 FAyA UNSE
2 CONICET
3 UPM España
4 EEA INTA Sgo del Estero
*Autor de contacto: albanesi@unse.edu.ar
Recibido: 18-04-15
Recibido con revisiones: 13-10-15
Aceptado: 13-10-15
RESUMEN
El objetivo de este trabajo fue evaluar el efecto del rolado de baja intensidad en indicadores biológicos y bioquímicos de calidad
del suelo, utilizado para habilitar un bosque secundario en un sistema silvopastoril (SSP) con siembra de Panicum maximum var. Gatton panic y sometido a pastoreo controlado (RBIs) en Santiago del Estero, Argentina bajo coberturas arbóreas de Ziziphus
mistol (M); Aspidosperma quebracho blanco (Qb), Schinopsis lorentzii (Qc) y suelo sin cobertura arbórea (D). Las variables
evaluadas fueron carbono orgánico total (COT), carbono orgánico particulado (COP), carbono de la biomasa microbiana
(Cbm), las relaciones COP: COT y Cbm:COT, glomalinas totales (GT) y fácilmente extraíbles (GFE). Para estudiar la relación
entre tratamientos, cobertura y variables de suelo se realizó ANOVA, test de Duncan para la comparación de medias (α =
0,05) y análisis de componentes principales (ACP). El SSP con siembra de gatton panic (RBIs) no modificó las variables evaluadas.
La cobertura arbórea modificó las variables evaluadas de tal manera que en áreas sin cobertura se registró una disminución
importante y aumentaron bajo cobertura arbórea de Qc, Qb y aún más de M. Los valores medios de la relación Cbm:COT
promediaron un 2% evidenciando que la biota del suelo tiene una baja eficiencia metabólica, propia de los ambientes
semiáridos.
Palabras clave: Biomasa microbiana; Región Chaqueña; Cobertura arbórea.
Silvopastoral system enable by a low intensity rolling with different tree
cover. Impact on glomalinas and soil carbon fractions
ABSTRACT
The aim of this study was to evaluate the effect of low intensity rolling on biological and biochemical indicators of soil
quality in a silvopastoral system (SSP) planted with Panicum maximum var. Gatton panic (RBIs) subjected to controlled
grazing, under tree covers (Ziziphus mistol (M); Aspidosperma quebracho blanco (Qb), Schinopsis lorentzii (Qc)) and a bare
soil (D) in Santiago del Estero, Argentina. We measure total organic carbon (TOC), particulate organic carbon (POC), microbial
biomass carbon (Cbm), POC:TOC and Cbm:TOC ratio, total glomalin (GT) and easily extractable glomalin (GFE) in bare soil,
without any vegetation cover (D) and soil under M; Qb and Qc canopies. ANOVA and a principal component analysis (PCA)
were performed in order to study the relationship between treatments, coverage and soil variables. The SSP planting Gatton
panic (RBIs) did not modify the evaluated variables. In areas without coverage, it was registered a significant decline in all
the evaluated variables and it was observed a significant increase of these variables under tree cover, especially under M
treatment. The mean value of CBM:COT relationship was 2% showing that soil biota has a low metabolic efficiency, typical
of semi-arid environments.
Key words: Microbial biomass; Chaco region; Tree cover.
INTRODUCCIÓN
La calidad del suelo es dinámica y puede cambiar en el
corto plazo de acuerdo con las características específicas
del mismo, las condiciones ambientales, el uso y las prácticas de manejo. Para conservarla es necesario implementar
diferentes prácticas sustentables de manejo según cada
situación particular (Navarrete Segueda et al., 2011).
En la Región Chaqueña la tala indiscriminada y el sobrepastoreo generaron grandes áreas con alta densidad de
leñosas espinosas y baja productividad de herbáceas para
la producción ganadera (Kunst et al., 2012).
Una de las prácticas sustentables que se proponen para
la habilitación de los bosques secundarios degradados es
la práctica del rolado de baja intensidad (RBI) para implementar sistemas silvopastoriles (Kunst et al., 2012) que
integran árboles y pasturas y generan ambientes más estables y diversos que los cultivos monofíticos y de mayor
productividad (Ramachandran et al., 2010). El rolado de baja
intensidad consiste en el pasaje del rolo de metal de bajo
peso con un mínimo número de pasadas que aplasta y corta
de forma selectiva el estrato arbustivo y deja el componente arbóreo brindando cobertura protectora al suelo, pasturas
y animales lo cual permite un mayor ingreso de luz y agua
que genera importantes aumentos en la biomasa herbácea y una mayor oferta y calidad de forraje de las especies
herbáceas nativas y más aún cuando se incorporan espe-
cies exóticas megatérmicas como Panicum maximum var
Gatton panic (Kunst et al., 2012). Debajo de la copa de los árboles existe un mayor rendimiento de materia seca de P. maximun y un mayor contenido de nutrientes especialmente C y N en suelos mientras que el pastoreo intensivo
tiene un fuerte efecto contrarrestando las influencias positivas de las plantas leñosas.
La importancia de las plantas leñosas en sistemas silvopastoriles de regiones semiáridas está relacionada a la
reducción de pérdida de agua y nutrientes (Silva et al., 2011;
Abule et al., 2005) y al mayor potencial para secuestrar
carbono orgánico que plantas herbáceas ya que los árboles contribuyen con una mayor proporción de C estabilizado (Lorenz & Lal, 2014).
El C orgánico del suelo (COS) es el principal componente de la materia orgánica y juega un rol trascendental
en el mantenimiento de la calidad de los suelos. Dentro de
la fracción de mayor labilidad, la fracción particulada constituida por material orgánico más joven y activo del suelo
y la biomasa microbiana juegan un papel muy importante
en el ciclo de los nutrientes (Galantini & Suñer, 2008).
La biomasa microbiana constituye entre el 1 y 3% del
C total del suelo y la disponibilidad de nutrientes y la productividad de los agroecosistemas dependen principalmente
del tamaño y de la actividad de la misma (Moore et al., 2000).
Dentro de la biomasa microbiana se encuentran entre
otros microorganismos, los hongos formadores de micorrizas arbusculares, que aportan carbono orgánico procedente de su masa miceliar extrarradical y producen glicoproteínas denominadas glomalinas las cuales contienen
un alto porcentaje de carbono orgánico (Rillig et al., 2003;
Schindler et al., 2007) que es incorporado directamente al
suelo mediante la descomposición de los propágulos
fúngicos (Driver et al., 2005). Las glomalinas se asocian
directamente con la estabilidad de los agregados del suelo
por su naturaleza insoluble y su alta resistencia a la descomposición (Rillig et al., 2001).
El rolado selectivo de baja intensidad con siembra de
pasturas (RBIs) modifica la estructura de la comunidad vegetal y por ende la producción primaria (Kunst et al., 2012).
Los cambios en la composición de la vegetación pueden
producir cambios sustanciales en la dinámica de los ciclos
biogeoquímicos ya que las especies vegetales y los factores abióticos ejercen influencia en las propiedades del suelo
determinando la calidad del mismo, especialmente en regiones áridas y semiáridas (Carranza et al., 2012).
El objetivo fue evaluar glomalinas, biomasa microbiana, carbono particulado y carbono orgánico total como
indicadores de calidad de la materia orgánica del suelo en
un sistema silvopastoril con cinco años de habilitación a
partir de un monte degradado mediante rolado de baja
intensidad y sometido a pastoreo controlado en Santiago
del Estero, Chaco Semiárido Argentino, bajo diferentes
coberturas arbóreas.
MATERIALES Y MÉTODOS
Área y Diseño Experimental
El estudio se realizó en el Campo Experimental ‘‘La María’’,
perteneciente a la Estación Experimental Agropecuaria del
Instituto Nacional de Tecnología Agropecuaria de Santiago
del Estero, Argentina (28º3´S y 64º15´O). El clima es semiárido
subtropical, con una precipitación anual promedio de 574 mm
y régimen monzónico. El suelo fue clasificado como Haplustol éntico, en el horizonte superficial (0-0,30 m) presenta pH 6,4,
densidad aparente de 0,9 g cm-3, textura franco limosa, estructura en bloques sub angulares y granular.
La vegetación es de bosque secundario caracterizado por
una comunidad vegetal dominada por especies arbustivas espinosas de géneros tales como Prosopis, Acacia, Celtis, Schinus, Larrea y especies arbóreas como Aspidosperma quebracho-blanco Schl., Schinopsis lorentzii Griseb., Prosopis spp., Zizyphus mistol Griseb (Kunst et al., 2012).
En el año 2006 se establecieron parcelas experimentales
de 2,5 ha cada una, completamente aleatorizadas con dos repeticiones por tratamiento, siendo:
T. Testigo: bosque secundario con una densidad de leñosas
mayor a 2000 tallos ha-1.
SSP. Sistema silvopastoril implementado con RBI: Dos
pasadas de rolo con un peso de 3000 kg a 45° con siembra simultánea de 5 kg ha-1 de Panicum maximum var Gatton panic
en 2006, más un pastoreo controlado por año (E.V. 1,1 ha-1)
en agosto de 2007, 2008, 2009, 2010 y 2011.
Como subtratamientos se estudió el efecto de la cobertura arbórea clasificada en: D. suelo sin cobertura arbórea; M.
suelo bajo cobertura de Ziziphus mistol (mistol); Qb. suelo bajo
cobertura de Aspidosperma quebracho blanco (quebracho
blanco) y Qc. suelo bajo cobertura de Schinopsis lorentzii
(quebracho colorado). Las especies arbóreas mencionadas
difieren en calidad y cantidad de residuos aportados así como
en período de senescencia.
En agosto de 2011 se tomaron 3 muestras compuestas de
suelo (n=3) a 15 cm de profundidad para cada cobertura (n=4)
y en cada tratamiento (n=2). Las muestras fueron secadas al
aire y tamizadas por tamiz de 2 mm.
Determinaciones analíticas
Carbono orgánico total (COT) (Nelson & Sommers, 1982),
determinado por espectrofotometría a 590 nm. Los resultados
se expresaron en g C kg-1 suelo.
Carbono orgánico particulado (COP) determinado en la
fracción de partículas comprendida entre los 2000 y 53 µm
separadas mediante tamizado en húmedo (Cambardella & Elliott, 1992) por espectrofotometría a 590 nm. Los resultados
se expresaron en g C kg-1 de suelo seco.
Se calculó la relación COP: COT, indicador indirecto del
efecto de diferentes manejos que modifican el aporte de carbono o su velocidad de descomposición (Galantini & Suñer, 2008).
Carbono de la biomasa microbiana (Cbm) por el método de
fumigación extracción (Vance et al., 1987) y espectrofotometría
a 590 nm. Los resultados se expresaron en mg C kg-1 de suelo.
Se calculó la relación Cbm:COT, que representa la proporción de COT contenido en la biomasa microbiana y se considera un indicador indirecto para monitorear cambios de la
materia orgánica en el suelo en la conversión de ecosistemas
naturales a sistemas agroforestales (Gomez et al., 2008).
Glomalinas totales, fracción más recalcitrante (GT), y fácilmente extraíbles, fracción más lábil débilmente unida a las
partículas del suelo (GFE) (Wright & Upadhyaya, 1996; González- Chavez et al., 2004). La extracción se realizó con citrato de sodio
50 mM en autoclave a 121°C por ciclos de 60 minutos y
centrifugación, repitiéndose la extracción hasta la desaparición
del color café-rojizo típico de la glomalina (se utilizó el primer
extracto para determinar GFE). La cuantificación se realizó por
espectrofotometría a 595 nm utilizando el método de Bradford
basado en el cambio de color del colorante Coomassie brilliant
blue G- 250 en respuesta a diferentes concentraciones de proteínas (Wright & Upadhyaya, 1996).
Análisis estadístico
Todas las variables fueron analizadas mediante ANOVA
cuyas fuentes de variación fueron tratamiento y cobertura,
previa verificación de los supuestos de normalidad y homogeneidad de varianza. Se utilizó test de Duncan para la comparación de medias (α = 0,05). Para estudiar la relación entre
las coberturas del SSP y las variables de suelo se realizó un
análisis de componentes principales (ACP) y de conglomerados. Todos los análisis se realizaron utilizando el paquete
estadístico InfoStat versión 2008p (Di Rienzo et al., 2008).
RESULTADOS Y DISCUSIÓN
Carbono orgánico total (COT), carbono‘particulado
(COP) y relación COP:COT
El COT y el COP no revelaron diferencias entre tratamientos y solo registraron diferencias significativas entre
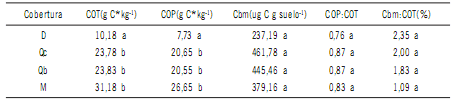
coberturas (p=0,016 y p= 0,022, respectivamente) (Tabla
1). En COT las magnitudes variaron entre 10,18 y 31,18
g C kg-1 y en COP entre 7,73 a 26,65 g C kg-1.
Tabla 1. Carbono orgánico total (COT), carbono particulado (COP), carbono de la biomasa microbiana (Cbm) y
relaciones COP:COT y Cbm:COT en suelo bajo cobertura de Schinopsis lorentzii (Qc), Aspidosperma quebracho-blanco (Qb), Zizyphus mistol (M) y sin cobertura (D).
Table 1. Total organic carbon (COT), particulate organic carbon (COP), microbial biomass carbon (Cbm) and
relationships COP:COT and Cbm:COT in soil under coverage Schinopsis lorentzii (Qc), aspidosperma quebracho-blanco (Qb ), Zizyphus mistol (M) and without cover (D).
Los menores valores medios de COT y COP se registraron en el suelo sin cobertura y los mayores valores en
suelo bajo cobertura arbórea. Estos resultados coinciden
con Hudak et al. (2003), quienes encontraron en otras
regiones semiáridas, mayores valores de COT en suelos
bajo cobertura de leñosas debido a que la vegetación contribuye en gran medida a la formación de materia orgánica
del suelo ya que constituye el principal material originario
de la misma mediante la formación de una capa de residuos orgánicos (mantillo), compuesta de hojas, ramas,
raíces, etc.
En los bosques del Chaco, la caída de hojarasca ocurre
a lo largo de todo el año. Sin embargo, la cantidad es baja
y varía según las características estructurales del bosque
(Montero et al., 2011) siendo superior en las áreas de dosel
cerrado con mayor abundancia de arbustos e inferior en
las áreas de dosel semi cerrado o abierto. Para bosques de
la región chaqueña húmeda se registró que el aporte por mantillo del quebracho colorado es de 380 kg ha-1 año
(Barberis et al., 2012) y no hay registro sobre el aporte del
Quebracho blanco y del Mistol.
El COT en este ambiente depende del aporte del mantillo y de la protección de la canopia por lo que la falta de
cobertura (D) se traduce en una disminución de COT por
la falta de aporte de mantillo y por el excesivo calentamiento, con la consecuente estimulación de la mineralización
del C, la formación de compuestos de más labilidad y la
menor protección a la degradación microbiana (Albanesi et al., 2003).
El mayor contenido medio de COP bajo cobertura
arbórea implica que existe más cantidad de C asociado a
macroagregados grandes y medianos más desprotegidos
de la acción microbiana (Cambardella & Elliot, 1992). Este
resultado indica la importancia de los aportes anuales y
consecutivos de materia seca aérea y radical que según
Galantini et al. (2008) implicarían una mayor contribución
al reciclado de nutrientes de las fracciones lábiles que
constituyen una fuente de energía y nutrientes de fácil
acceso para los microorganismos saprófitos responsables
de dicha transformación.
El menor valor medio de COP se registró en D ya que
el incremento de COP está en relación directa con los
aportes anuales de los residuos de la vegetación por lo que
la ausencia de cobertura vegetal disminuye el C en superficie y afecta la dinámica del COP (Álvarez & Steinbach,
2006).
La relación COP:COT no registró diferencias estadísticas significativas entre tratamiento, cobertura ni en la
interacción tratamiento*cobertura y varió entre 76 a 87%,
evidenciando que la mayor cantidad de C está fácilmente
disponible para la degradación microbiana, coincidiendo con García Oliva et al. (1999) quienes indicaron que en suelos
forestales el 80% del C está asociado al COP. En este trabajo el carbono orgánico asociado a limos y arcillas (como
resultado de la diferencia entre COT y COP) varió entre
2,45 a 4,53 g C kg-1 es decir que la materia orgánica humificada o que forma un complejo órgano mineral es escasa,
por lo que se debería evitar la rápida pérdida del COP para
el mantenimiento del sistema (Videla et al., 2008).
Carbono de la Biomasa Microbiana (Cbm)
y relación Cbm:COT
El Cbm no registró diferencias significativas en ninguna fuente de variación considerada. Se observaron mayores valores bajo cobertura arbórea y menores en D, evidenciando que el manejo de la cobertura del suelo influye en
los microorganismos y los procesos microbianos del suelo
por la cantidad y calidad de los residuos vegetales aportados, la estacionalidad del aporte y la distribución espacial
(Christensen et al., 1994; Kandeler et al., 1999).
La relación Cbm:COT no registró diferencias estadísticas significativas entre tratamientos, cobertura ni en la
interacción tratamiento*cobertura. Los resultados indican
que el SSP y las coberturas no modifican la eficiencia metabólica de la biota del suelo, es decir la eficiencia de los microorganismos para formar biomasa a expensas del COT.
Los valores medios de la relación Cbm:COT promedió en
un 2% (Tabla 1). Estos valores se encontraron dentro de
los valores informados en la bibliografía (Moore et al.,
2000), que señala que alrededor del 1 al 3% del carbono
total del suelo es biomasa microbiana. La biota de estos
suelos presenta una baja eficiencia metabólica, por lo que
refleja también una baja contribución de la biomasa microbiana al C orgánico del suelo, propio de los ambientes
semiáridos.
Glomalinas
Las glomalinas totales (GT) y fácilmente extraíbles
(GFE) no registraron diferencias significativas entre tratamientos, lo que evidencia que el SSP obtenido con RBIs no
afectó a las mismas. Esto podría deberse al bajo impacto
de la práctica sobre el componente arbóreo concordando
con Driver et al. (2005) quienes reportan que los niveles
de glomalinas serían superiores en ecosistemas forestales
que en agroecosistemas, posiblemente debido al incremento en la velocidad de descomposición de los diversos
propágulos de los hongos micorrícicos arbusculares (HMA)
presentes en los suelos bajo bosque, proceso responsable
de la incorporación de las glomalinas a la fracción húmica
del suelo.
Las glomalinas totales (GT) y fácilmente extraíbles
(GFE) registraron diferencias significativas entre coberturas (p= 0,0028 y 0,0016, respectivamente). Los mayores
valores medios de glomalinas se registraron en suelos bajo
cobertura de M y los menores valores medios se registraron en D (Fig. 1), en oposición a Burrows (2014) quien
reportó que la diversidad de plantas está más vinculada a
la producción de glomalinas que la cobertura vegetal.
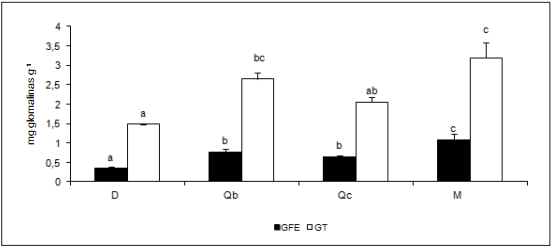
Figura 1. Valores medios de glomalina fácilmente extraíble (GFE), y glomalina total (GT) expresadas en mg de glomalina g-1 suelo bajo cobertura de Schinopsis lorentzii (Qc), Aspidosperma quebracho-blanco (Qb), Zizyphus mistol (M) y en suelo sin cobertura (D). Letras diferentes indican diferencias
estadísticas significativas (Duncan α = 0,05).
Figure 1. Mean values of easily removable Glomalin (GFE) and total Glomalin (GT) expressed in mg g-1 soil under coverage Schinopsis lorentzii (Qc), Aspidosperma
quebracho-blanco (Qb), Zizyphus mistol (M) and bare soil (D). Different letters indicate statistically significant differences (Duncan α = 0.05).
Los resultados observados coinciden con Rillig et al.
(2003), quienes indicaron que los suelos cubiertos con
especies arbóreas o pastos presentan mayor concentración
de glomalinas que los suelos libres de vegetación, lo cual explica la presencia permanente de plantas que propician
abastecimiento permanente de fotosintatos a los hongos
micorrícicos arbusculares (HMA) estrechamente relacionados con la producción de glomalinas (González-Chávez et al., 2004).
Los mayores valores medios de GT y GFE se observaron bajo la cobertura de M, ya que esta especie aporta mayor
cantidad de residuos orgánicos con menor relación C:N y
mayor cantidad de mucílagos, taninos, almidón y azúcares
que Qc y Qb (Colares & Arambarri, 2008), de fácil aprovechamiento para los hongos micorrícicos.
Rillig et al. (2002) indican que la composición de la
comunidad de plantas puede influir sobre la concentración
de glomalinas en el suelo. Las raíces sirven como sitios
hospederos de los HMA, pero el hospedante influye diferencialmente sobre las especies de HMA y tiene efecto
directo sobre el suelo. La zona de mayor actividad radical en
Qc se registra en el horizonte A del suelo (profundidad de
0,10 m) y existe una rápida absorción y redistribución del
fósforo en la planta (Barberis et al., 2012).El tipo de sistema radical de las especies arbóreas puede influir en la
producción de glomalinas de los HMA, pudiendo ser el de
M el más favorable en este sentido.
Los individuos de especies leñosas (Qc, Qb y M) realizan un aporte regular de residuos orgánicos y cobertura
vegetal permanente, en comparación a sistemas productivos de pasturas a cielo abierto, favoreciendo la conservación de la humedad, la actividad biológica, la proliferación
de HMA y la producción de glomalinas tal como reportan
Borie et al. (2006). No hay información acerca de interacciones de Schinopsis lorentzii con hongos micorrícicos aún
cuando se registró en Schinopsis haenkeana y Schinopsis brasiliensis la presencia de micorrizas arbusculares con colonización tipo Arum-Paris (Barberis et al., 2012).
Análisis de Conglomerados y Componentes
principales
En el análisis de conglomerados (Fig. 2), según la distancia euclídea (medida de similaridad) se agruparon los
tratamientos y coberturas fijando un criterio de corte en
la distancia 2,39 (distancia igual al 50% de la distancia máxima, criterio de corte frecuentemente utilizado), seleccionada con el fin de agrupar las variables con máxima homogeneidad en cada grupo y con mayor diferencia entre
los mismos. Aun cuando el proceso de agrupamiento
conlleva inicialmente a una pérdida de información, la síntesis de la información disponible puede facilitar la visualización de relaciones multivariadas (Di Rienzo et al., 2008).
Con este criterio los tratamientos bajo cobertura de M (T:M
y SSP:M) se separaron del resto al encontrarse más distantes. Los tratamientos bajo cobertura de Qb y Qc formaron
otro grupo y el tercer grupo estuvo constituido por los
tratamientos sin cobertura (TD y SSPD).
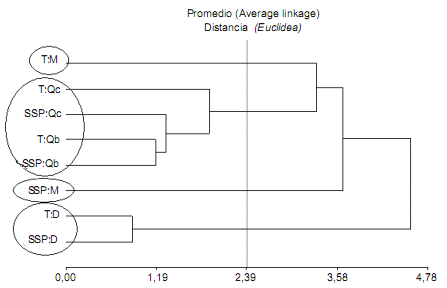
Figura 2. Análisis de Conglomerados. Referencias: Tratamientos: Testigo (T), sistema silvopastoril implementado con RBI (SSP). Coberturas: Schinopsis lorentzii (Qc), Aspidosperma quebracho-blanco (Qb), Zizyphus mistol (M) y suelo sin cobertura (D).
Figure 2. Cluster Analysis. References: treatments: control (T), RBI implemented silvopastoral system (SSP). Coverage: Schinopsis lorentzii (Qc), Aspidosperma quebracho-blanco (Qb), Zizyphus mistol (M) and bare soil (D).
Los primeros componentes principales (CP 1 y CP 2)
explicaron el 94% de la variabilidad (Fig. 3) y las variables
con mayor peso, es decir las que presentaron mayores
valores de auto-vectores siendo éstos coeficientes que
ponderan las variables y muestran la inercia relativa de cada
una de ellas en la variabilidad total. Además de relacionar
las variables, explican el efecto de los tratamientos y la cobertura sobre la calidad del suelo. Dentro de la CP 1 las variables de mayor peso fueron COP, COT, GT y GFE, con
valores de auto-vectores de 0,45, 0,44, 0,43, 0,43, respectivamente, y en la CP 2 fueron Cbm:COT y Cbm con valores de autovectores de 0,67 y 0,59 respectivamente. Las
CP correlacionaron positivamente con las variables originales siendo la correlación de la CP 1 con COP, COT, GT
y GFE de 0,99, 0,98, 0,94, 0,96, respectivamente y CP 2
con Cbm:COT y Cbm de, 0,77 y 0,62, respectivamente.
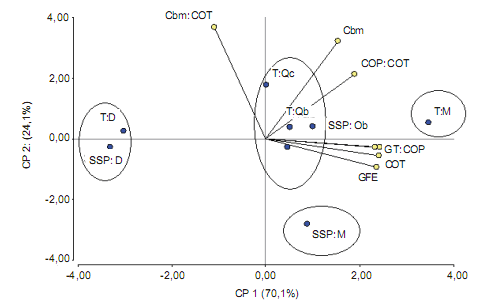
Figura 3. Análisis biplot de los tratamientos, coberturas y variables de suelo. Referencias: Tratamientos: Testigo (T), sistema silvopastoril implementado
con RBI (SSP). Coberturas: Schinopsis lorentzii (Qc), Aspidosperma quebracho-blanco (Qb), Zizyphus mistol (M) y suelo sin cobertura (D). Variables
de suelo: carbono orgánico total (COT), carbono orgánico particulado (COP), carbono de la biomasa microbiana (Cbm), glomalina fácilmente extraíble
(GFE), glomalina total (GT) y relaciones COP:COT y Cbm:COT
Figure 3. Analysis biplot treatments, covers and soil variables. References: treatments: control (T), RBI implemented silvopastoral system (SSP).
Coverage: Schinopsis lorentzii (Qc), Aspidosperma quebracho-blanco (Qb), Zizyphus mistol (M) and bare soil (D). Soil variables: total organic carbon
(COT), particulate organic carbon (COP), microbial biomass carbon (Cbm), readily removable Glomalin (GFE) Glomalin Total (GT) and relationships COP:
COT and Cbm: COT
Los ángulos formados entre los vectores que representan a las variables GFE y GT estuvieron correlacionados
positivamente con las variables químicas (COT y COP)
(Fig.3). Esto concuerda con Seguel et al. (2008) quienes encontraron una alta relación positiva entre las glomalinas
y la materia orgánica del suelo en estudios de bosques nativos
en Chile. Los suelos cubiertos con especies arbóreas o pastos presentan mayor concentración de glomalinas que los
suelos libres de vegetación, lo cual se explica por la presencia
permanente de plantas que propician abastecimiento permanente de fotosintatos para alimentar los hongos micorrícicos arbusculares estrechamente relacionados con la
producción de glomalinas (Gonzalez-Chavez et al., 2004;
Rilling et al., 2003).
En el plano bidimensional construido a partir de las dos
primeras componentes principales se distinguieron claramente cuatro grupos (Fig. 3): el primero constituido por los
tratamientos sin cobertura (T:D, SSP:D), el segundo por los tratamientos bajo cobertura de quebrachos (T:Qc, T:Qb, SSP:Qc
y SSP:Qb), el tercero y cuarto por tratamientos bajo cobertura de mistol (T:M y SSP:M). El primer grupo al encontrarse a la izquierda de la CP 1 se caracterizó por menores
contenidos de COT, COP, GT y GFE; el segundo se caracterizó por valores intermedios y el tercero y cuarto grupo, al encontrarse a la derecha de la CP 1, se caracterizaron
por magnitudes mayores de dichas variables (Fig. 3).
Estos resultados posiblemente se deban a que la cantidad de proteínas y carbohidratos solubles en hojas de mistol
es elevada (Nogués et al., 2013), por lo que la incorporación
de las mismas como residuos al suelo aportarán al COT del
suelo, especialmente a las fracciones más lábiles.
El tratamiento T:Qc se caracterizó por mayor Cbm y
relación Cbm:COT ya que se encontró en la parte superior
de la CP 2, en oposición a SSP:M, indicando en este último
caso diferencias en la calidad de la materia orgánica o a su
disponibilidad por los microorganismos (Insam & Domsch,
1988; Guerrero et al., 2003), los cuales presentan baja
eficiencia metabólica.
Estos resultados indican que se debe seguir monitoreando los cambios en el tiempo ya que estas variables
biológicas y microbiológicas responden más rápidamente a los cambios en el suelo que las variables químicas y podrían evidenciar el efecto del SSP en menor tiempo, a pesar
que en el presente trabajo no se encontraron diferencias
significativas en ninguna de ellas debido a que en la conversión de bosques nativos en sistemas silvopastoriles
semiáridos, la permanencia de los árboles mantiene el
contenido de nutrientes y materia orgánica mientras que
el reemplazo completo del bosque a pastura implantada
como, por ejemplo sucedió en ecosistemas semiáridos de
Brasil, disminuyó el contenido de Cbm aun cuando no
modificó las reservas de nutrientes y materia orgánica (Wick et al., 2000).
CONCLUSIONES
El SSP implementado mediante RBIs no modifica las
propiedades químicas y microbiológicas en el suelo. La cobertura arbórea tiene un efecto significativo en los contenidos de glomalinas, biomasa microbiana, carbono particulado y carbono orgánico total. Estas variables son
mayores bajo cobertura arbórea de Quebracho colorado
y blanco y aún más de Mistol. La biota del suelo presenta
una baja eficiencia metabólica, propia de los ambientes
semiáridos. Se debe continuar monitoreando en el tiempo
para adquirir información certera del efecto del pastoreo.
AGRADECIMIENTOS
Consejo de Investigaciones en Ciencia y Técnica de la
Universidad Nacional de Santiago del Estero.
Instituto Nacional de Tecnología Agropecuaria EEA
Santiago del Estero.
BIBLIOGRAFÍA
1. Abule, E; GN Smit & HA Snyman. 2005. The influence of woody plants
and livestock grazing on grass species composition, yield and soil
nutrients in the Middle Awash Valley of Ethiopia. Journal of Arid
Environments 60: 343-358.
2. Albanesi, A; A Anriquez & A Polo Sánchez. 2003. Efectos de la agricultura convencional en algunas formas del C en una toposecuencia
de la Región Chaqueña, Argentina. Agriscientia Vol 20: 9-20.
3. Álvarez, R & HS Steinbach. 2006. Factores climáticos y edáficos
reguladores del nivel de materia orgánica. En: Materia Orgánica «Valor Agronómico y Dinámica en Suelos Pampeanos» (ed. R Álvarez). Editorial. Facultad de Agronomía, Universidad de Buenos
Aires. p. 31-40.
4. Barberis, IM; V Mogni; L Oakley; C Alzugaray; JL Vesprini & De Prado.
Biología de especies australes : Schinopsis balansae Engl. (Anacardiaceae). Kurtziana [online]. 2012, vol.37, n.2 [citado 2013-12-17],
pp. 59-86. Disponible en: <http://www.scielo.org.ar/scielo.php?
script=sci_arttext&pid=S1852-59622012000200006 &lng=es&nrm=iso>. ISSN 1852-5962.
5. Borie, F; R Rubio; J Rouanet; A Morales; G Borie & C Rojas. 2006. Effect
of tillage systems on soil characteristics, glomalin and mycorrhizal
propagules in a Chilean Ultisol. Soil & Tillage Res. 88: 253-261.
6. Burrows, RL. 2014. Glomalin Production and Infectivity of Arbuscular-Mycorrhizal Fungi in Response to Grassland Plant Diversity.
American Journal of Plant Sciences 5, 103-111 Published Online
January 2014 (http://www.scirp.org/journal/ajps) http://dx.doi.org/10.4236/ajps.2014.51013
7. Cambardella, C & E Elliott. 1992. Particulate soil organic-matter
changes across a grassland cultivation sequence. Soil Sci. Soc. Am. J. 56: 777-783.
8. Carranza, C; L Noe; C Merlo; M Ledesma & A Abril. 2012. Efecto del
tipo de desmonte sobre la descomposición de pastos nativos e
introducidos en el Chaco Árido de la Argentina. RIA Vol. 38 Nº 1,
97-107.
9. Christensen, NB; WC Lindemann; E Salazar-Sosa & LR Gill. 1994.
Nitrogen and carbon dynamics in no-till and stubble mulch tillage
systems. Agron. J. 86, 298-303.
10. Colares, MN & AM Arambarri. 2008. Ziziphus mistol (Rhamnaceae):
Morfo-anatomía y arquitectura foliar. Latin American Journal of
Pharmacy (formerly Acta Farmacéutica Bonaerence). Lat. Am. J.
Pharm. 27 (4): 568-77.
11. Di Rienzo, JA; F Casanoves; MG Balzarini; L Gonzalez; M Tablada & CW Robledo. 2008. InfoStat, versión 2008, Grupo InfoStat, FCA,
Universidad Nacional de Córdoba, Argentina.
12. Driver, J; W Holben & M Rillig. 2005. Characterization of glomalin
as a hyphal wall component of arbuscular mycorrhizal fungi. SoilBiol. Biochem. 37: 101-106
13. Galantini, J; J Iglesias; M Landriscini; L Suñer & G Minolo. 2008. Calidad
y dinámica de las fracciones orgánicas en sistemas naturales
cultivados. En: Galantini J. (ed). Estudio de las fracciones orgánicas
en suelos de la Argentina. 1º edición. Editorial de la Universidad
Nacional del Sur. Bahía Blanca. Argentina. 308 p.
14. Galantini, JA & L Suñer. 2008. Las fracciones orgánicas del suelo: análisis
en los suelos de la Argentina. Agricientia Vol XXV (1): 41-55.
15. García-Oliva, F; Jr R Sanford & E Kelly. 1999. Effects of slash and burn
management on soil Aggregate Organic C and N in a Tropical
Deciduous Forest. Geoderma 88:1-12.
16. Gomez Y; J Paolini & RM Hernández. 2008. Sustitución de la sabana
nativa con plantaciones de Pinus cariaea (Pinaceae) en Venezuela:
efecto sobre parámetros indicadores de cambios en el carbono del
suelo. Rev. Biol. Trop. (Int. J. Trop. Biol.) 56(4): 2041-2053.
17. González-Chávez, MCA; MC Gutiérrez-Castorena & S Wright. 2004.
Hongos micorrízicos arbusculares en la agregación del suelo y su
estabilidad. TERRA Latinoamericana Vol. 22, Núm. 4 pp. 507-514.
18. Guerrero, C; J Mataix-Solera; F Rodríguez; F García-Orenes; I Gómez & R Moral. 2003. Carbono microbiano edáfico en suelos afectados
por el fuego y enmendados con diferentes residuos orgánicos. Edafología 10(2): 185-190.
19. Hudak, AT; CA Wessman & TR Seastedt. 2003. Woody overstorey
effects on soil carbon and nitrogen pools in South African savanna. Austral Ecology 28: 173-181.
20. Insam, H & KH Domsch. 1988. Relationship between soil organic
carbon and microbial biomass on chronosequences of reclamation
sites. Microbial Ecol. 15: 177-188.
21. Kandeler, E; D Tscherko & H Spiegel. 1999. Long-term monitoring
of microbial biomass, N mineralization and enzyme activities of
a Chernozem under different tillage management. Biol. Fert. Soils
28, 343-351.
22. Kunst, C; R Ledesma; S Bravo; A Albanesi; A Anriquez; H Van Meer & J Godoy. 2012. Disrupting woody steady states in the Chaco
region (Argentina): Responses to combined disturbance treatments. Ecological Engineering 42: 42-53.
23. Lorenz, K & R Lal. 2014. Soil organic carbón sequestration in agroforestry systems. A review. Agron. Sustain. Dev. 34: 443-454.
24. Montero, GA; NJ Carnevale & G Magra. 2011. Ensambles estacionales
de artrópodos epigeos en un bosque de quebracho (Schinopsis balansae) en el Chaco Húmedo. Revista Colombiana de Entomología 37(2): 294-304.
25. Moore, JM; S Klose & MA Tabatabai. 2000. Soil microbial biomass
carbon and nitrógeno as affected by cropping Systems. Biol Fertil
Soils 31: 200-210.
26. Navarrete Segueda, A; G Vela Correa; J López Blanco & ML Rodríguez
Gamiño. 2011. Naturaleza y utilidad de los indicadores de calidad
del suelo. Contactos 80: 29-37.
27. Nelson, D & L Sommers. 1982. Methods of soil analysis. In: Page,
A. (ed). Agronomy 9, ASA, SSSA. 539-579.
28. Nogués, EM; OE Castro; RJ Correa; M Puricelli; H Gómez Pérez & R López.
2013. Ziziphus mistol Griseb. (mistol) su rol en la religión, la cultura
popular, la alimentación humana y animal y su uso medicinal en
el árido y semiárido del NOA. Información general sobre la importancia de este género en distintas regiones del mundo. Revista de
Divulgación Técnica Agrícola y Agroindustrial Facultad de Ciencias Agrarias-UNCa. 52: 1-16.
29. Ramachandran Nair, PK; VD Nair; B Mohan Kumar & JM Showalter.
2010. Carbon sequestration in agroforestry systems. In: D. Sparks (ed) Advances in Agronomy, Vol. 108 1° ed Pp, 237-307, Academic
Press. USA, 326 p.
30. Rillig, M; S Wright; K Nichols; W Shmith & M Torn. 2001. Large
contributions of arbuscular mycorrhizal fungi to soil carbon pools
in tropical forest soils. Plant Soil 233: 167-177.
31. Rillig, MC; SF Wright & VT Eviner. 2002. The role of arbuscular
mycorrhizal fungi and glomalin in soil aggregation: Comparing
effects of five plant species. Plant Soil 238: 325-333.
32. Rillig, M; P Ramsey; S Morris & E Paul. 2003. Glomalin, an arbuscularmycorrhizal fungal soil protein, responds to soil-use change. PlantSoil 253(2): 293-299.
33. Seguel, A; R Rubio; R Carrillo; A Espinosa & F Borie. 2008. Niveles
de glomalina y su relación con características químicas y biológicas del suelo (andisol) en un relicto de bosque nativo del sur de
Chile. Bosque 29(1): 11-22.
34. Schindler, F; E Mercer & J Rice. 2007. Chemical characteristics of
glomalin-relatad soil protein (GRSP) extracted from soils of varying
organic matter content. Soil Biol. Biochem. 39: 320-329.
35. Silva, GL; HV Lima; MM Campanha; RJ Gilkes & TS Oliveira. 2011
Soil physical quality of Luvisols under agroforestry, natural
vegetation and conventional crop management systems in the
Brazilian semi-arid region. Geoderma 167-168: 61-70.
36. Vance, ED; PC Brookes & DS Jenkinson. 1987. An extraction method
for measuring soil microbial biomass C. Soil Biol. Biochem. 19: 703-707.
37. Videla, LS; CM Rostagno & MA Toyos. 2008. La materia orgánica
particulada: comparación de métodos para su determinación y su
valor como indicador de calidad de suelos del Chubut. Revista Ciencia
del Suelo (Argentina) 26(2): 219-227.
38. Wick, B; H Tiessen & RSC Menezes. 2000. Land quality changes
following the conversion of the natural vegetation into silvo-pastoral systems in semi-arid NE Brazil. Plant and Soil 222: 59-70.
39. Wright, SF & A Upadhyaya. 1996. Extraction of an abundant and
unusual protein from soil and comparison with hyphal protein of
arbuscular mycorrhizal fungi. Soil Sci. 161: 575-586.